
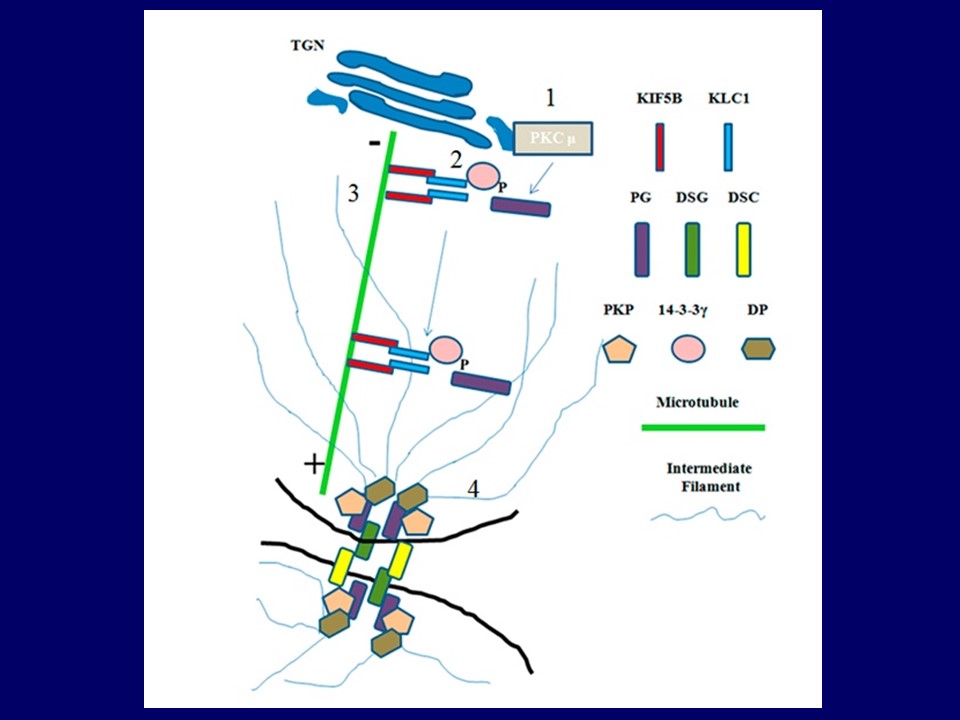
Work in the laboratory has led to the development of a novel method for the generation of transgenic animals. US and Indian patent applications for the process have been filed and the protocol has been published. Further, our work has shown that loss of 14-3-3γ leads to sterility in male mice due to a defect in desmosome formation between sertoli cells and also between Sertoli cells and spermatocytes. The loss of adhesion seems to be due to a decrease in desomosome formation, due to a defect in the transport of the desmosomal plaque protein, plakoglobin (PG), to the cell border. This leads to defects in Blood:Testis barrier formation, inhibiting spermatocyte differentiation. Our work has also demonstrated that this is probably due to the inability of 14-3-3γ to load the cargo PG onto the kinesin motor protein, KIF5B. Loss of either KIF5B or 14-3-3γ in the testis leads to a disruption of cell-cell adhesion and sterility. We are now focused on determining whether 14-3-3γ is required for desmosome formation in other tissues and cell types.
To determine the role of 14-3-3γ in regulating desmosome formation in vitro, we hypothesized that it was possible that the only function of 14-3-3γ in regulating the desmosome formation was in transporting plakoglobin (PG) to the cell border as we have reported. If this was the case then if we could artificially localize PG to the cell border in cells lacking 14-3-3γ, this should induce the formation of the desmosome. To test this hypothesis, a farnesylated version of PG fused to GFP was introduced into 14-3-3γ knockdown cells. It was observed that in the transfected cells, desmosomal proteins, desmoplakin (DP), desmocollin 2/3 (DSC2/3) and plakophilin3 (PKP3), re-localized to the border in contrast to cells transfected with a farnesylated GFP protein (GFP-f), even though both the PG-GFP-f and GFP-f localize to the cell border. These results suggested that the main role of 14-3-3γ in regulating desmosome formation is to transport PG to the border to induce desmosome formation.
Loss of 14-3-3ε leads to a lymphoproliferative disorder leading to splenomegaly and lymphocyte infiltration in the lungs, liver and kidney leading to death of the animal. In addition to the splenomegaly, the mice also show hair loss with a decrease in hair formation and alterations in the organization of the epidermis. We are now attempting to generate tissue specific knockdown mice for 14-3-3ε to study epidermal differentiation and hair follicle development in vivo. This project has just been initiated in the laboratory and we are in the process of generating the mice required to test these hypotheses